血液腫瘤科/細胞治療中心/台灣細胞免疫醫學會 陳駿逸醫師
雌激素受體訊號傳導
雌激素受體(ER)分為核受體和膜受體。
ERα 和 ERβ 是核受體,能夠抑制或活化目標基因。兩種受體均具有 6 個功能域,並具有多種相似性,且能夠形成異二聚體。
ERα 在乳癌的進展和發病機制中發揮重要作用,因為其在乳癌中表達上調。最新研究表明,約 75% 的乳癌患者 ERα 表達呈陽性。
值得注意的是,ERβ 抑制可活化乳癌 1 型(BRCA1)基因,進而抑制腫瘤細胞增生。然而,ERβ 的表達會隨著乳癌細胞增殖的增加而降低。值得注意的是,ERβ 在小鼠模型中透過與 p53 相互作用發揮抑癌作用。
相反,ERα 透過與細胞週期蛋白 D1 相互作用促進乳癌細胞的進展。細胞週期蛋白 D1 在某些腫瘤細胞中調節細胞週期從 G1 期到 S 期的轉換。因此,ERα與細胞週期蛋白D1的相互作用被認為是ERα促進乳癌細胞增生的最重要機制。
根據結構,ERα可分為多種亞型,其中ERα36亞型可促進轉移並加重乳癌患者的病情。相反,ERα之Tanixufen亞型是乳癌治療的標靶,它透過上調醛脫氫酶1家族成員A1 (ALDH1A1)基因,對ERα36具有拮抗作用。
表觀遺傳修飾
組蛋白修飾
組蛋白修飾,例如:乙醯化(乙醯輔酶A的乙醯基與賴氨酸氨基共價結合)、去乙醯化(去除組蛋白賴氨酸尾部的乙醯基)和甲基化(在組蛋白賴氨酸和精胺酸尾部添加甲基),在乳癌中的作用已被廣泛研究,如下圖所示。組蛋白去乙醯化(簡稱HDAC)會導致染色質結構更加緊密,並抑制組蛋白基因的表達。此外,乳腺組織中正常細胞和癌細胞的組蛋白乙醯化和甲基化指標也存在差異。

圖示顯示,癌症基因和抑癌基因的失調是由於CpG島甲基化和組蛋白改變所致。
此外,已鑑定出用於區分乳癌亞型的標記物。例如,管腔A型和基底型乳癌亞型均表現出H3K4me3、H3K27me3和H3K27me3的下調。然而,H3K27me3的過度表現僅在管腔A型中檢測到,這啟發了乳癌亞型內聚類的概念。
值得注意的是,在患者來源的癌組織、原發性乳癌細胞和乳癌細胞系中評估了3個H3賴氨酸乙醯化、14個H3賴氨酸甲基化以及14個賴氨酸組合變異的組蛋白改變。組蛋白修飾功能異常(例如H3K9ac/H3K27me3失衡)對癌症、腫瘤細胞增殖和增殖、乳癌訊號通路以及轉移的影響已被廣泛研究。此外,研究顯示H3K9ac不穩定性與細胞分裂、凋亡、細胞間通訊、遷移和能量代謝相關的基因有關聯。
組蛋白去乙醯化酶(HDAC)依其與酵母蛋白的相似性分為四類。 HDAC調控多種參與乳癌發生發展的蛋白質的表達和產生。在乳癌中已觀察到HDAC的過度表現。基質金屬蛋白酶 9 (MM9 蛋白編碼基因) 和增強的細胞侵襲與 BC 細胞中 HDAC1、HDAC8 或 HDAC6 的過度表現有關。
DNA甲基化
在DNA甲基化過程中,S-腺苷甲硫胺酸(SAM)上的甲基可逆地轉移到胞嘧啶的第五位碳原子上,此反應由DNA甲基轉移酶(DNMTs)催化,例如:DNMT3A、DNMT1和DNMT3B。然而,這些DNMTs的功能各不相同。例如,DNMT3A和DNMT3B在胚胎髮生過程中形成新的DNA甲基化模式,而DNMT1僅參與將甲基添加到DNA上。
全基因組定序顯示乳癌的發生與DNA甲基化模式之間存在關聯,並在40種不同的乳腺腫瘤細胞系中鑑定出345個甲基化基因。此外,對目標CpG位點的進一步分析發現了超過18700個差異甲基化基因。而且,在乳癌中也發現了基因體低甲基化現象。值得注意的是,基因體內的CpG甲基化可能促進轉錄激活,而啟動子區域的CpG甲基化則與基因表現呈負相關。
此外,DNA甲基化影響多個參與轉錄訊號傳導、DNA結合、同源異型盒蛋白和細胞分化的基因。 DNA甲基化也影響染色質結構、轉錄調控、DNA修復、細胞週期調控、細胞凋亡和代謝活動,所有這些都與惡性細胞的發生發展密切相關。此外,參與細胞黏附、組織侵襲和惡性腫瘤路徑的基因也受到DNA甲基化變異的影響。更重要的是,能夠影響腫瘤微環境的DNA甲基化可能改變腫瘤發生過程。
微小RNA
無論癌症處於哪個階段,微小RNA(miRNA)都與乳房腫瘤的發病機制密切相關。 miRNA具有致癌和抑癌的雙重作用,在癌症中扮演雙重角色。在所檢測的309種miRNA中,有133種在各種乳癌亞型的健康和惡性組織中均扮演重要角色。此外,miRNA還可用於乳癌的分期。值得注意的是,miRNA可以調控雌激素受體(ER)的表達。總體而言,miRNA在正常乳房發育和乳癌(BC)發展中起著至關重要的作用。然而,某些miRNA,例如miRNA-27a,則作為致癌因子,促進ER過度表達,從而促進癌症發展。
微小RNA在乳癌中的作用
miRNA與乳癌的原因密切相關,既可以發揮抑制癌的作用,也可以發揮促癌作用。有些miRNA可以調控ER的表達,並可用於乳癌的分期。 miRNA,例如microRNA-335-5p、microRNA-21、microRNA-142-3p和microRNA-192-5p,能夠抑制ER表達;而另一些miRNA,例如miRNA-27a,則能夠促進ER過表達,從而促進癌症發展。在MCF-7細胞中,miRNA-148a透過降低DNMT1的表達來上調ER表達。總之,miRNA在正常乳腺形成和乳腺腫瘤組織的發生和發展中都扮演著至關重要的角色。
在乳癌的發生與發展過程中,miRNA參與細胞生長、上皮間質轉化(EMT)、能量代謝、Wnt/β-catenin訊號路徑、免疫反應和細胞凋亡等過程。相反,microRNA-1207-5p則透過長鏈非編碼RNA PVT1促進細胞增殖。包括microRNA-543、microRNA-455、microRNA-143、microRNA-424和microRNA-26a-5p在內的多種microRNA能夠抑制細胞增殖。由於SerpinB2和miR-200c/141簇能夠調控參與EMT和轉移的基因,因此被認為是三陰性乳癌的預測指標。
在惡性腫瘤患者中觀察到microRNA-200c、microRNA-455和microRNA-141的表達上調。 microRNA-122透過增強營養物質向轉移前部位的可及性來促進轉移,而miR-148則抑制乳癌細胞的進展和擴散。
此外,microRNA也參與能量代謝。包括microRNA-421、microRNA-193a、microRNA-133a、microRNA-211-5p、microRNA-335、microRNA-145和microRNA-124在內的多種miRNA能夠抑制腫瘤擴散<sup>105,106</sup>。 CD274簇的3’-UTR可被microRNA(如microRNA-195和microRNA-497)靶向,從而促進腫瘤細胞的免疫逃脫並抑制免疫反應。此外,miRNA還有助於腫瘤細胞逃避凋亡,這至關重要。
環狀RNA在乳癌中的作用
研究證據表明,環狀RNA(circRNA)參與乳癌、黑色素瘤的發生和擴散。此外,乳房組織中的circRNA含量低於正常組織,而鄰近正常組織樣本中的circRNA可能作為乳癌的指標。 circRNA晶片分析鑑定出乳癌患者中差異表達的circRNA,包括上調的circRNA hsa、circ100219、hsa circ406697和circ006054,以及下調的circRNA circ103110、circ104689和circ104821。
此外,某些產生大量circRNA的蛋白質編碼等位基因與腫瘤相關。目前,研究人員正在努力闡明circRNA在黑色素瘤中的功能。
環狀RNA在基因調控中的作用
大多數生物活動,包括癌症的發生和發展,都依賴基因表現的調控。環狀RNA(circRNA)可作為microRNA海綿,調控癌細胞的增生、轉移與遷移。例如,circHIPK3可吸附9種miRNA,包括miR-124,後者是參與乳腺腫瘤組織發育的腫瘤抑制因子。此外,在乳癌細胞中敲低hsa-circ 0001982可抑制細胞侵襲和增殖,並誘導細胞凋亡。類似地,敲低circ-ABCB10可透過吸附miR-1271促進乳癌細胞凋亡並抑制其增殖。
circGFRA1可促進細胞增生並抑制細胞凋亡,且與腫瘤大小、TNM分期、淋巴結轉移和三陰性乳癌的組織學分級密切相關。 circGFRA1可能作為microRNA-34a海綿,透過競爭性內源性RNA(ceRNA)機制調控GFRA1的表現。綜上所述,這些發現表明,circRNA可以透過捕獲與腫瘤發生、增殖、分化和遷移相關的特定miRNA來調控癌症的發生與發展。
環狀RNA在治療中的作用
近期研究發現,某些環狀RNA(circRNA)對乳癌的發生與發展至關重要,並可能成為潛在的治療標靶。例如,circRNA-000911透過吸附microRNA-449a,促進Notch 1或核因子κB(NF-κB)訊號路徑的活性;而circ-FOXO3則能誘導細胞凋亡,減緩乳癌的擴散。 CircVRK1抑制乳癌幹細胞(BCSC)的擴散和自我更新能力,使其成為降低乳癌轉移和復發的潛在治療標靶。清除乳癌幹細胞對於減少乳房黑色素瘤的擴散和復發至關重要。總而言之,自從發現circRNA參與乳癌的生長和發展以來,人們已經開展了大量研究來闡明其作為潛在治療標靶的作用。
乳癌放射抗藥性和粒線體氧化壓力的表觀遺傳調控
放射治療仍然是乳癌治療的關鍵組成部分,尤其是在局部治療和術後治療。然而,放射抗性的產生顯著限制了其長期療效。表觀遺傳修飾已成為放射抗性的關鍵介質,其作用機制通常是透過改變DNA修復能力、染色質重塑以及非編碼RNA表達失調來實現的。 DNMTs和HDACs在抗藥性腫瘤中普遍過度表達,導致抑癌基因沉默和放射誘導的細胞凋亡受損。
此外,粒線體在氧化壓力調控和表觀遺傳訊號傳導中都扮演核心角色。放射線治療會誘導粒線體功能障礙,導致活性氧(ROS)水平升高,加劇基因組的不穩定並促進腫瘤細胞的存活。粒線體代謝產物,例如:α-酮戊二酸和乙醯輔酶A,會影響核和粒線體的表觀遺傳標記,從而調節參與壓力反應和能量代謝的基因表現。
研究表明,粒線體DNA甲基化模式和組蛋白乙醯化的改變會影響氧化磷酸化基因的轉錄,從而促進治療抗藥性。靶向氧化還原敏感的轉錄因子和粒線體表觀遺傳的調控因子有望提高抗藥性腫瘤對放射治療的敏感性。因此,整合粒線體和氧化壓力驅動的表觀遺傳機制的知識對於理解放射抗性以及開發更有效的乳癌治療策略至關重要。
腫瘤微環境要素
乳癌細胞可轉移至骨骼、肝臟、肺臟、腦部等組織,在轉移組織中,固有基質細胞與擴散的腫瘤細胞之間會發生多種交互作用。生長因子、免疫細胞、趨化因子、細胞外基質(ECM)、細胞激素、腫瘤相關巨噬細胞、腫瘤相關纖維母細胞等尚未確定的成分均參與腫瘤微環境的形成。器官特異性因素和各種基質細胞的侵襲均可以影響轉移性的微環境。
固有基質細胞與擴散的腫瘤細胞在各種宿主組織中的相互作用錯綜複雜。多種因素可能影響轉移性微環境,尤其是在器官和各種基質細胞。
免疫系統細胞
免疫系統抑制腫瘤復發和腫瘤發生。除了對感染的免疫反應外,白血球在癌症的發生與擴散中也扮演著重要角色。遠處腫瘤組織可藉由改變亞臨床白血球組成而導致轉移。中性粒細胞是一類白血球,可能在轉移性疾病的啟動中發揮作用。
據報道,在乳癌小鼠模型中,特定類型的嗜中性白血球,例如 :CXCR2+ 嗜中性球,會影響轉移後間質細胞的活性。對乳癌小鼠模型中轉移前肺微環境的分析表明,嗜中性白血球作為轉移啟動的調節因子發揮作用,這表明免疫細胞控制著惡性微環境的建立。具體而言,在黑色素瘤中,CXCR2+嗜中性球可誘導間質基質細胞的促轉移作用。此外,研究人員透過改變乳癌小鼠模型中的癌前肺部微環境,闡明了嗜中性白血球在轉移發生中的作用,這表明免疫系統細胞可能控制惡性微環境的形成。
值得注意的是,免疫系統對腫瘤發生既有抑制作用,也有促進作用,這取決於癌症的類型和分期。例如,遠離初始腫瘤部位的白血球含量改變可導致轉移,這使得白血球成為促進腫瘤生長和轉移的關鍵免疫細胞。
轉化生長因子β(TGF-β)
TGF-β透過調節多種發炎細胞因子(S100A8/A9)和生長因子(例如:血管內皮生長因子A (VEGF) 和血管生成素2 (ANGPT2))來輔助建構轉移前的肺部微環境。 TGF-β的異常產生會透過改變微環境促進乳癌的進展。例如,TGF-1受體激酶 (ALK5) 抑制劑IN-1130在4T1黑色素瘤原位小鼠模型中具有預防肺轉移的潛力。此外,另一種ALK-5抑制劑EW-7197可預防乳癌患者的肺轉移。基於這些發現,可以推測,單獨或合併免疫療法抑制TGF-β訊號路徑可能是治療黑色素瘤引起的肺轉移的潛在方法。
TGF-β異常表現所引起的微環境改變可能影響黑色素瘤的發生與發展。值得注意的是,利用4T1異種移植模型,研究人員闡明了TGF-β影響特定細胞激素(S100A8/A9)和生長因子(VEGF、Angpt2)從而促進轉移前微環境形成的機制。綜上所述,這些發現提示,免疫療法合併抑制TGF-β訊號路徑可能是治療乳房癌肺轉移的有效策略。
細胞外基質蛋白
在乳癌侵襲的早期階段,細胞外基質(ECM)蛋白,例如:versican、tenascin-C (TNC) 和periostin (POSTN),對於轉移病灶的形成至關重要。乳癌幹細胞會表現TNC,而TNC通常由纖維母細胞產生。此外,乳癌幹細胞中TNC的異常表現在肺轉移中具有啟動作用。
另一種可以與Wnt配體結合的基質來源成分是POSTN。研究表明,POSTN在肺癌發展過程中能夠促進癌幹細胞的生長。同樣,浸潤的CD11b+/Ly6C髓系細胞在肺部惡性部位產生versican,促進肺轉移。此外,ECM成分可能透過為癌細胞在體內擴散提供適宜的微環境來促進腫瘤轉移的發展。自然殺手細胞、巨噬細胞和其他免疫細胞表達整合素41,與纖維連接蛋白相連,並與血管細胞黏附分子-1 (VCAM-1) 結合。值得注意的是,VCAM-1在乳癌細胞中異常表現。表達VCAM-1的黑色素瘤傾向歸巢於肺組織實質,因為肺組織實質含有膠原蛋白和彈性蛋白纖維。
乳癌基因的突變
同源定向DNA修復促進BRCA蛋白的協同作用,從而減少和預防腫瘤。因此,BRCA基因的缺失突變或損傷可能會降低DNA修復能力,進而增加癌症風險。例如,BRCA基因的缺失突變和功能喪失會使乳腺腫瘤的風險增加5至6倍,增加惡性細胞增殖的可能性。
BRCA1和BRCA2分別有1650種和1731種不同的突變。 BRCA1和BRCA2基因中最常見的突變是移碼突變,這種突變通常會導致提前出現終止密碼子,並降低成熟RNA和功能性蛋白質的水平。研究結果表明,年輕的乳癌患者,尤其是病情較重的乳癌患者,BRCA突變的比例更高。此外,臨床證據表明,基底上皮標記的表達與攜帶 BRCA1 突變但 ERBB2 或 ER 未過度表現的乳腺腫瘤之間存在關聯。
這些指標通常與 ERBB2 或 ER未表達的腫瘤相關。儘管這些癌細胞表現出基底樣表型,但 BRCA-1 基底樣乳腺黑色素瘤細胞和自發性基底樣乳癌起源於管腔上皮前體細胞,而非基底祖細胞。值得注意的是,腫瘤表型可能並不總是反映具有表型變異的細胞的組織學特徵。乳癌經常會活化 PI3K/AKT/雷帕黴素標靶蛋白 (mTOR) 通路,該通路被認為是一種致癌通路。
大約30%的情況下,PIK3CA改變和磷酸酶及張力蛋白同源物(PTEN)蛋白缺失是最常見的PI3K活化路徑。自2004年在多種實體瘤中發現以來,編碼p110a的PIK3CA體細胞突變已被通報。甚至在PIK3CA突變被發現之前,PIK3CA基因過度表現已在多種腫瘤中觀察到,包括約10%的乳癌病例。然而,僅約3%的癌症存在PIK3R1突變。整體而言,PIK3R1基因產物p85a透過維持p110a發揮負向之腫瘤調控作用。
值得注意的是,在表達ER和PR的乳腺腫瘤中,AKT1突變(E17K)的發生率介於1.4%至8%之間。如果ER陽性腫瘤發生改變,激活PI3K級聯反應,例如PTEN下調、HER2或類胰島素生長因子1受體(IGF-1R)上調,或突變AKT1激活,則可能對荷爾蒙治療產生抗藥性。有效的預後生物標記,例如PIK3CA突變,可以幫助乳癌患者的臨床治療。
幹細胞在乳癌中的作用
幹細胞在乳癌的發生發展和治療抗藥性的各個方面都發揮著重要作用。癌症幹細胞是腫瘤內的一個細胞亞群,具有自我更新和分化為腫瘤內各種細胞類型的能力。人們認為癌症幹細胞是腫瘤起始和維持的關鍵因素。
在乳癌中,癌症幹細胞導致腫瘤異質性,因為它們可以分化為侵襲性不同的細胞類型。癌症幹細胞通常對化療和放療等傳統癌症治療方法具有抗藥性,導致腫瘤在初始治療後復發。癌幹細胞的固有特性,包括其外排泵活性,使其能夠存活並抵抗治療作用。癌症幹細胞參與腫瘤向體內遠端部位的轉移過程。此外,癌症幹細胞在轉移性腫瘤的擴散和起始中起著至關重要的作用,促進乳癌擴散到其他器官。
腫瘤微環境,包括血管、免疫細胞和支持細胞,與癌幹細胞相互作用。微環境訊號可以影響癌症幹細胞的行為,包括自我更新、分化和侵襲。幹細胞(包括癌症幹細胞)可以發生上皮間質轉化(EMT),從而增強癌細胞侵襲周圍組織和轉移至身體其他部位的能力。目前,針對癌症幹細胞的標靶治療策略正被廣泛研究,旨在有效治療乳癌。標靶阻斷癌症幹細胞自我更新和存活相關路徑可望改善治療效果。
了解幹細胞在乳癌中的作用有助於開發個人化治療方案,從而提高治療效果。此外,識別並針對個別腫瘤內的癌症幹細胞,有望開發出針對特定癌症特徵的客製化治療方案。幹細胞,尤其是癌症幹細胞,在乳癌的發生與發展、治療抗藥性和轉移過程中發揮著至關重要的作用。
結語
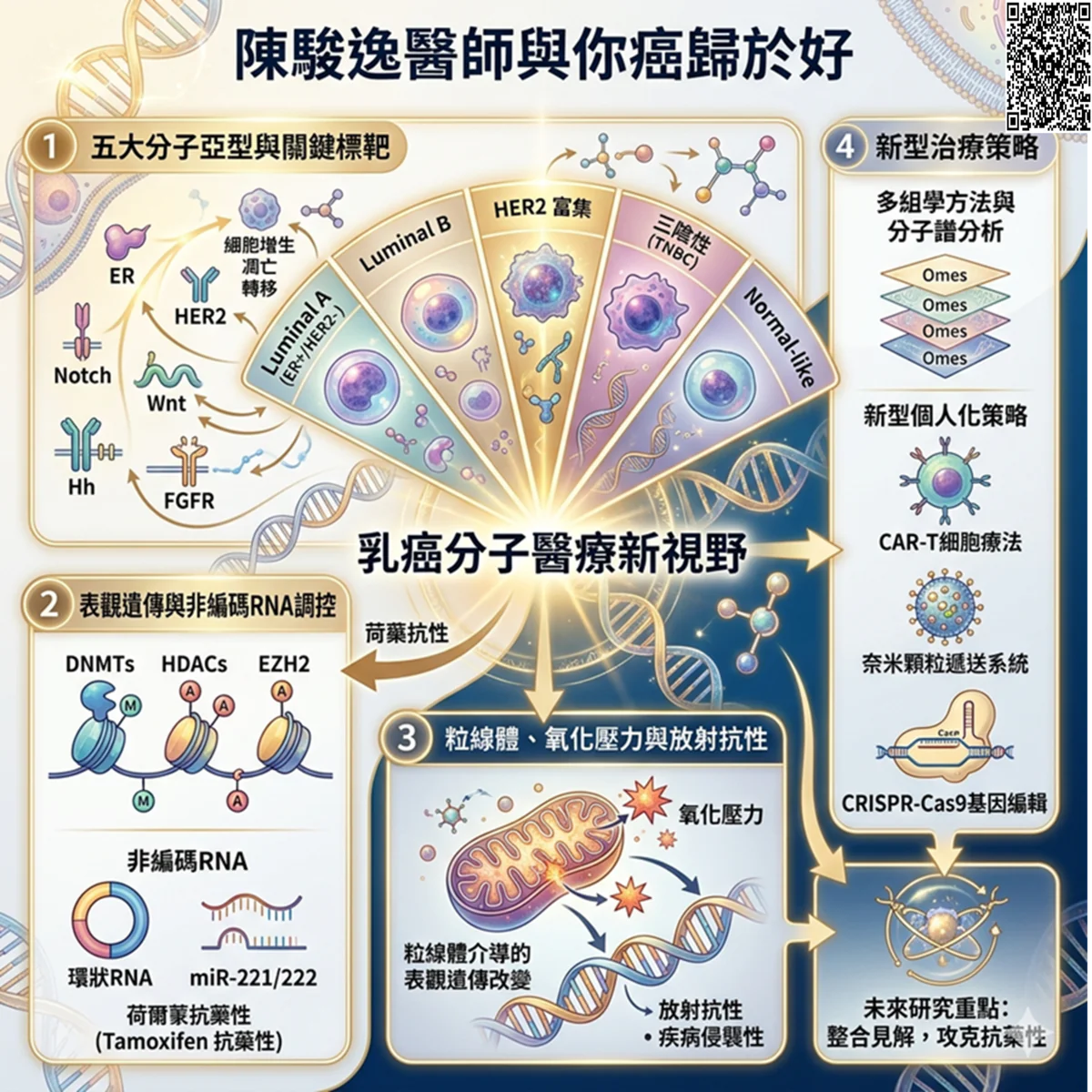
乳癌目前在分子層面上被分為5個亞型,每個亞型都有獨特的基因表現和訊號路徑特徵。其中最有前景的分子標靶包括ER、HER2、Notch、Wnt、Hh和FGFR,它們調控細胞增生、凋亡和轉移等關鍵過程。
此外,包括DNMTs、HDACs、增強子組蛋白同源物2 (EZH2) 在內的表觀遺傳調控因子以及miR-221/222和環狀RNA等非編碼RNA與荷爾蒙抗藥性密切相關,尤其是Tamoxifen抗藥性乳癌。
粒線體介導的表觀遺傳學改變和氧化壓力進一步加劇了放射抗性和疾病侵襲性。多組學方法和分子譜分析的進步已促成了預測性生物標記和治療標靶的發現。
CAR-T細胞療法、基於奈米顆粒的遞送系統和CRISPR-Cas9基因編輯等新型策略為個人化醫療開闢了新的方向。未來的研究應著重整合這些分子層面的見解,以開發針對抗藥性機制的標靶之干預措施,並改善乳癌患者的臨床結果。
醫學重點解析與整合
一、 雌激素受體(ER)的分子拉鋸戰
- ERα 的促癌機制:ERα 在約 75% 的乳癌患者中呈現陽性表達。它主要透過與細胞週期蛋白 D1 (Cyclin D1) 相互作用,加速細胞週期從 G1 期轉換至 S 期,這是促進乳癌細胞增生最關鍵的機制。其中,ERα36 亞型會進一步促進腫瘤轉移並惡化病情。
- ERβ 的抑癌角色:與 ERα 相反,ERβ 透過與 p53 蛋白相互作用發揮抑癌活性;激活 ERβ 可活化 BRCA1 基因進而抑制細胞增生。然而,隨著乳癌細胞惡性增殖,ERβ 的表達量會逐漸降低。
二、 表觀遺傳學與 DNA 甲基化的雙重調控
- 組蛋白修飾異常:組蛋白去乙醯化(HDAC)會使染色質結構緊密,抑制抑癌基因表達。例如 HDAC1、HDAC6 或 HDAC8 的過度表現,會直接上調基質金屬蛋白酶 9(MM9),增強乳癌細胞的侵襲能力。
- DNA 甲基化失衡:由 DNMT1、DNMT3A 和 DNMT3B 催化的甲基化轉移具有位置特異性。啟動子區域的 CpG 島甲基化與基因表現呈負相關(會導致基因沉默);而全基因組的低甲基化與特定的表觀變異,則會引發染色質結構不穩定,進而激活惡性轉錄訊號。
三、 非編碼 RNA(miRNA 與 circRNA)的致癌與抑癌網絡
- 微小 RNA (miRNA) 的雙向調節:
- 抑制 ER 表達:microRNA-335-5p, -21, -142-3p, -192-5p。
- 促進 ER 表達:miRNA-27a 作為致癌因子促進 ER 過度表達;miRNA-148a 則透過降低 DNMT1 來間接上調 ER。
- 免疫逃逸:microRNA-195 和 -497 靶向 CD274 簇的 3′-UTR,助長腫瘤細胞逃避免疫反應。
- 環狀 RNA (circRNA) 的「miRNA 海綿」機制:
- circHIPK3 與 circ-ABCB10 可分別吸附抑制因子(如 miR-124、miR-1271),調控癌細胞的增殖與凋亡。
- circGFRA1 在三陰性乳癌中異常關鍵,可作為 microRNA-34a 的海綿以維持 GFRA1 的表達,其表達水平與 TNM 分期及淋巴結轉移密切相關。
四、 粒線體氧化壓力與放射抗藥性
- 放療耐受機制:DNMTs 和 HDACs 在抗藥性腫瘤中普遍過度表達,導致放射誘導的細胞凋亡路徑受阻。
- 粒線體逆向訊號:放射治療引發的粒線體功能障礙會導致活性氧(ROS)積聚。粒線體代謝產物(如 $\alpha$-酮戊二酸、乙醯輔酶A)會改變核內的表觀遺傳標記,進而惡化能量代謝並誘導對放療的抗性。
五、 腫瘤微環境(TME)與癌症幹細胞(CSCs)
- 微環境屏障:免疫細胞(如 CXCR2+ 嗜中性球)、TGF-β(透過調控 VEGF 和 ANGPT2 構築轉移前靶器官微環境)以及細胞外基質蛋白(如 TNC、POSTN)相互交織。乳癌幹細胞自身會分泌 TNC 來啟動肺轉移。
- 癌症幹細胞的威脅:CSCs 具有無限自我更新與分化能力,其高表達的外排泵活性是導致化療與放療全面失效、驅動上皮間質轉化(EMT)及引發遠端復發的根本原因。
#雌激素受體
#ERα
#ERβ
#BRCA1
#表觀遺傳修飾
#組蛋白乙醯化
#甲基化
#HDAC
#DNMTs
#CpG島超甲基化
#非編碼 RNA
#miRNA-27a
#miR-148a
#circRNA
#circGFRA1
#陳駿逸醫師
#台中市全方位癌症關懷協會
陳駿逸醫師醫療門診服務時段
https://mycancerfree.com/contact/
更多陳駿逸醫師的癌症衛教影片請連接https://www.youtube.com/@mycancerfree
更多腫瘤治療相關資訊 請連接”陳駿逸醫師 與你癌歸於好” https://mycancerfree.com